
| Overview | Cell Structures | Cell Migration | Cell Division |
Control of the Location of the Rear Rather than Front by Centrosome |
Zhang et al., Mol. Biol. Cell. 28:3240-3251 (2018) |
Centrosome is often located in front of the nucleus relative to the direction of migration, which led to the conventional view that centrosome may dictate the position of the front. However there are many exceptions to this observation. We have proposed that microtubules may be responsible for the global distribution of inhibitory signals, away from the front to create a tail |
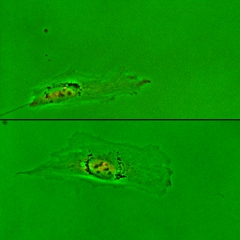 |
Rear Localization of Centrosome in Cells Undergoing Persistent Migration |
Two representative RPE-1 cells expressing GFP-centrin (a centrosomal protein; red dots indicated by white arrowheads) are migrating on unconfined 2D surfaces. The top cell is migrating persistently from left to right, with its centrosome localized to the rear. The bottom cell is migrating randomly, and its centrosome is located close to the center. Images are taken every 4 min for 332 min. |
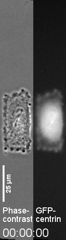
Positioning of Centrosome to the Future Rear before the Onset of Directional Migration |
|
A freshly plated RPE-1 cell expressing GFP-centrin (a centrosomal protein; red dots indicated by white arrowheads) is initially spreading in both directions along a micropatterned adhesive linear strip, while centrosome is located closer to the lower end. This end subsequently becomes the tail after the cell starts directional migration. Images are taken every 6 min for 102 min. |
 |
Positioning of Centrosome to the Future Rear before the Onset of Directional Migration |
Similar to the video above, with more footage during the spreading process. An RPE-1 cell expressing GFP-centrin (as indicated by red arrowheads) is spreading along a micropatterned adhesive linear strip after adhesion. The centrosome fluctuates around the cell centroid. The cell breaks symmetry after the centrosome becomes more stably located toward one end, which then becomes the retracting tail. Images are taken every 6 min for 258 min. |
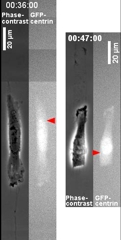 |
Redirection of Cell Migration by Micromanipulation of Centrosome Position |
Two RPE-1 cells expressing GFP-centrin (as indicated by red arrowheads) are cut at both ends, which causes transient spreading initially. In the left cell, the centrosome remains closer to the top end of the cell, which becomes the tail as the cell resumes directional migration. In the right cell, the centrosome moves across the nucleus to the other side of the cell after cutting, which becomes the tail as the cell resumes directional migration. Video starts after the second cut for both cells, images for the left cell are then taken every 6 min until 186 min after the second cut, and images for the right cell are taken every 4 min until 115 min after the second cut. |
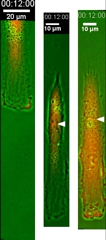 |
Movement of Centrosome Prior to Directional Reversal |
Three RPE-1 cells expressing GFP-centrin (red dots as indicated by white arrowheads) are migrating along a vertical strip before hitting a dead end at the bottom. The centrosome moves downward after the cell hits the deadend, and continues even after new protrusions appear at the opposite end. The cell does not reverse the direction until the centrosome has moved sufficiently close to the lower end prior to its retraction to form a tail. Images are taken every 6 min for 456 min (left cell), 192 min (middle cell), and 324 min (right cell), respectively. |
 |
Persistent Migration of Centrosome-Containing Cytoplasts |
Cells are treated with cytochalasin B followed by centrifugation to remove the nucleus. Previous studies indicate that the great majority of the resulting cytoplasts contain centrosome. Persistent migration is observed in both control and enucleated RPE-1 (upper), NIH3T3 (middle), and CEF (lower) cells along 10-μm-wide strips. All cells are stained with Hoechst (colored in red) to determine if they have been enucleated or not. Images are taken every 4 min for 11 h. |
 |
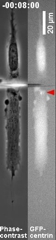
Persistent Migration of Centrosome-Containing Cell Fragments Created by Microsurgery |
An RPE-1 cell expressing GFP-centrin (as indicated by red arrowheads) is cut by a microneedle between the nucleus and the centrosome so that the fragment under observation contains only the centrosome. The fragment subsequently develops a persistent polarity. Images are taken every 4 min until 249 min after the microsurgery. |
 |
Constriction of the Cell around the Centrosome to Form a Pseudo Tail in Cell Fragments |
An RPE-1 cell expressing GFP-centrin (as indicated by red arrowheads) is cut by a microneedle between the nucleus and the centrosome so that the fragment contains only the centrosome. The fragment undergoes severe constriction around the centrosome to form a tail like region, while both ends extend in opposite directions. As a result of the competition between the two ends, the fragment is unable to migrate directionally. Images are taken every 6 min until 274 min after the microsurgery. |
 |
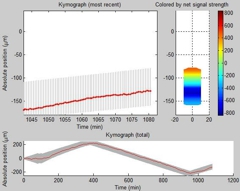
Dependence of Tail Formation and Persistent Migration on Centrosome |
On the left side of the video, an RPE-1 cell expressing GFP-centrin (as indicated by red arrowheads) is cut so that the fragment under observation only contains the nucleus but no centrosome. Although the cell manages to migrate slowly upward, it exhibits a defect in tail retraction. On the right side of the video, another RPE-1 cell expressing GFP-centrin is cut so that the fragment under observation contains neither a nucleus nor a centrosome. The fragment is unable to undergo directional migration. Images are taken every 6 min until 693 min (left cell) and 577 min (right cell) after the microsurgery, respectively. |
 |
Computer Simulation of the Role of Centrosome during Symmetry Breaking, |
Animation of a model cell, colored with the heat map of protrusion/retraction signals is shown in the top-right panel. The model is described in a separate video |